процесс анаэробного расщепления органических веществ, преимущественно углеводов, происходящий под влиянием микроорганизмов или выделенных из них ферментов (См.
Ферменты). В ходе Б. в результате сопряженных окислительно-восстановительных реакций освобождается энергия, необходимая для жизнедеятельности микроорганизмов, и образуются химические соединения, которые микроорганизмы используют для биосинтеза аминокислот, белков, органических кислот, жиров и др. компонентов тела. Одновременно накапливаются конечные продукты Б. В зависимости от их характера различают Б. спиртовое, молочнокислое, маслянокислое, пропионовокислое, ацетоно-бутиловое, ацетоно-этиловое и др. виды. Характер Б., его интенсивность, количественные соотношения конечных продуктов, а также направление Б. зависят от особенностей его возбудителя и условий, при которых Б. протекает (pH, аэрация, субстрат и др.).
Спиртовое Б. В 1836 французский учёный Каньяр де ла Тур установил, что спиртовое Б. связано с ростом и размножением дрожжей. Химическое уравнение спиртового Б.: C6H12O6 → 2C2H5OH + 2CO2 было дано французскими химиками А. Лавуазье (1789) и Ж. Гей-Люссаком (1815). Л. Пастер пришёл к выводу (1857), что спиртовое Б. могут вызывать только живые дрожжи в анаэробных условиях ("брожение - это жизнь без воздуха"). В противовес этому немецкий учёный Ю. Либих упорно настаивал на том, что Б. происходит вне живой клетки. На возможность бесклеточного спиртового Б. впервые (1871) указала русский врач-биохимик М. М. Манассеина. Немецкий химик Э. Бухнер в 1897, отжав под большим давлением дрожжи, растёртые с кварцевым песком, получил бесклеточный сок, сбраживающий сахар с образованием спирта и CO2. При нагревании до 50°C и выше сок утрачивал бродильные свойства. Всё это указывало на ферментативную природу активного начала, содержащегося в дрожжевом соке. Русский химик Л. А. Иванов обнаружил (1905), что добавленные к дрожжевому соку фосфаты в несколько раз повышают скорость Б. Исследования отечественных биохимиков А. И. Лебедева, С. П. Костычева, Я. О. Парнаса и немецких биохимиков К. Нейберга, Г. Эмбдена, О. Мейергофа и др. подтвердили, что фосфорная кислота участвует в важнейших этапах спиртового Б.
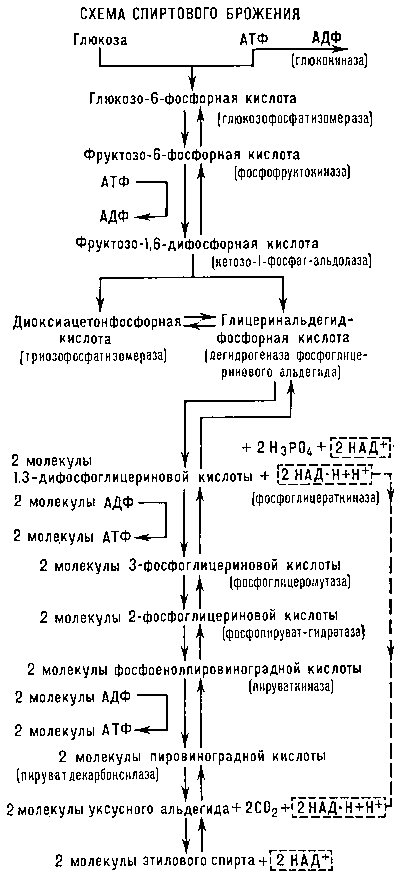
В дальнейшем многие исследователи детально изучили ферментативную природу и механизм спиртового Б. (см. схему). Первая реакция превращения глюкозы при спиртовом Б. - присоединение к глюкозе под влиянием фермента глюкокиназы остатка фосфорной кислоты от аденозинтрифосфорной кислоты (АТФ, см.
Аденозинфосфорные кислоты). При этом образуются аденозиндифосфорная кислота (АДФ) и глюкозо-6-фосфорная кислотата. Последняя под действием фермента глюкозофосфати-зомеразы превращается в фруктозо-6-фосфорную кислоту, которая, получая от новой молекулы АТФ (при участии фермента фосфофруктокиназы) ещё один остаток фосфорной кислоты, превращается в фруктозо-1,6-дифосфорную кислоту. (Эта и следующая реакции, обозначенные встречными стрелками, обратимы, т. е. их направление зависит от условий - концентрации фермента, pH и др.) Под влиянием фермента кетозо-1-фосфатальдолазы фруктозо-1,6-дифосфорная кислота расщепляется на глицеринальдегидфосфорную и диоксиацетонфосфорную кислоты которые могут превращаться друг в друга под действием фермента триозофосфатизомеразы. Глицеринальдегидфосфорная кислота, присоединяя молекулу неорганической фосфорной кислоты и окисляясь под действием фермента дегидрогеназы фосфоглицеринальдегида, активной группой которого у дрожжей является никотинамидадениндинуклеотид (НАД), превращается в 1,3-дифосфоглицериновую кислоту. Молекула диоксиацетонфосфорной кислоты под действием триозофосфатизомеразы даёт вторую молекулу глицеринальдегидфосфорной кислоты, также подвергающуюся окислению до 1,3-дифосфоглицериновой кислоты; последняя, отдавая АДФ (под действием фермента фосфоглицераткиназы) один остаток фосфорной кислоты, превращается в З-фосфоглицериновую кислоту, которая под действием фермента фосфоглицеро-мутазы превращается в 2-фосфоглицериновую кислоту, а она под влиянием фермента фосфопируват-гидратазы - в фосфоенол-пировиноградную кислоту. Последняя при участии фермента пируваткиназы передаёт остаток фосфорной кислоты молекуле АДФ, в результате чего образуется молекула АТФ и молекула енолпировиноградной кислоты, которая весьма нестойка и переходит в пировиноградную кислоту. Эта кислота при участии имеющегося в дрожжах фермента пируватдекарбоксилазы расщепляется на уксусный альдегид и двуокись углерода. Уксусный альдегид, реагируя с образовавшейся при окислении глицеринальдегидфосфорной кислоты восстановленной формой никотинамидадениндинуклеотида (НАД-Н), при участии фермента алкогольдегидрогеназы превращается в этиловый спирт. Суммарно уравнение спиртового Б. может быть представлено в следующем виде:
C6H12O6 + 2H3PO4 + 2АДФ → 2CH3CH2OH + 2CO2 + 2АТФ.
Т. о., при сбраживании 1
моля глюкозы образуются 2
моля этилового спирта, 2
моля CO
2, а также в результате фосфорилирования 2
молей АДФ образуются 2
моля АТФ. Термодинамические расчёты показывают, что при спиртовом Б. превращение 1
моля глюкозы может сопровождаться уменьшением свободной энергии примерно на 210
кдж (50 000
кал), т. е
. энергия, аккумулированная в 1
моле этилового спирта, на 210
кдж (50 000
кал) меньше энергии 1
моля глюкозы. При образовании 1
моля АТФ (макроэргических - богатых энергией фосфатных соединений) используется 42
кдж (10 000
кал). Следовательно, значительная часть энергии, освобождающейся при спиртовом Б., запасается в виде АТФ, обеспечивающей разнообразные энергетические потребности дрожжевых клеток. Такое же биологическое значение имеет процесс Б. и у др. микроорганизмов. При полном сгорании 1
моля глюкозы (с образованием CO
2 и H
2O) изменение свободной энергии достигает 2,87
Мдж (686 000
кал). Иначе говоря, дрожжевая клетка использует лишь 7\% энергии глюкозы. Это показывает малую эффективность анаэробных процессов по сравнению с процессами, идущими в присутствии кислорода. При наличии кислорода спиртовое Б. угнетается или прекращается и дрожжи получают энергию для жизнедеятельности в процессе дыхания. Наблюдается тесная связь между Б. и дыханием микроорганизмов, растений и животных. Ферменты, участвующие в спиртовом Б., имеются также в тканях животных и растений. Во многих случаях первые этапы расщепления сахаров, вплоть до образования пировиноградной кислоты, - общие для Б. и дыхания. Большее значение процесс анаэробного распада глюкозы имеет и при сокращении мышц (см.
Гликолиз), первые этапы этого процесса также сходны с начальными реакциями спиртового Б.
Сбраживание углеводов (глюкозы, ферментативных гидролизатов крахмала, кислотных гидролизатов древесины) используется во многих отраслях промышленности: для получения этилового спирта, глицерина и др. технических и пищевых продуктов. На спиртовом Б. основаны приготовление теста в хлебопекарной промышленности, виноделие и пивоварение.
Молочнокислое Б. Молочнокислые бактерии подразделяют на 2 группы - гомоферментативные и гетероферментативные. Гомоферментативные бактерии (например, Lactobacillus delbrückii) расщепляют моносахариды с образованием двух молекул молочной кислоты в соответствии с суммарным уравнением:
C6H12O6 = 2CH3CHOH·COOH.
Гетероферментативные бактерии (например, Bacterium lactis aerogenes) ведут сбраживание с образованием молочной кислоты, уксусной кислоты, этилового спирта и CO2, а также образуют небольшое количество ароматических. веществ - диацетила, эфиров и т.д.
При молочнокислом Б. превращение углеводов, особенно на первых этапах, близко к реакциям спиртового Б., за исключением декарбоксилирования пировиноградной кислоты, которая восстанавливается до молочной кислоты за счёт водорода, получаемого от НАД-Н. Гомоферментативное молочнокислое Б. используется для получения молочной кислоты, при изготовлении различных кислых молочных продуктов, хлеба и в силосовании кормов в сельском хозяйстве. Гетероферментативное молочнокислое Б. происходит при консервировании различных плодов и овощей путём квашения.
Маслянокислое Б. Сбраживание углеводов с преимущественным образованием масляной кислоты производят многие анаэробные бактерии, относящиеся к роду Clostridium. Первые этапы расщепления углеводов при маслянокислом Б. аналогичны соответстветственным этапам спиртового Б., вплоть до образования пировиноградной кислоты, из которой при маслянокислом Б. образуется ацетил-кофермент A (CH3CO-KoA). Ацетил-KoA может служить предшественником масляной кислоты, подвергаясь следующим превращениям:
Маслянокислое Б. применялось для получения масляной кислоты из крахмала.
Ацетоно-бутиловое Б. бактерии Clostridium acetobutylicum сбраживают углеводы с преим. образованием бутилового спирта (CH3CH2CH2CH2OH) и ацетона (CH3COCH3). При этом образуются также в сравнительно небольших количествах водород, CO2, уксусная, масляная кислоты, этиловый спирт. Первые этапы расщепления углеводов те же, что и при спиртовом Б. Бутиловый спирт образуется путём восстановления масляной кислоты:
CH3CH2CH2COOH + 4H = CH3CH2CH2CH2OH + H2O.
Ацетон же образуется декарбоксилированием ацетоуксусной кислоты, которая получается в результате конденсации двух молекул уксусной кислоты. Исследованиями В. Н. Шапошникова показано, что ацетоно-бутиловое Б. (как и ряд др., например пропионовокислое, маслянокислое) в опытах с растущей культурой происходит в две фазы. В первую фазу Б. параллельно с нарастанием биомассы накапливаются уксусная и масляная кислоты; во вторую фазу образуются преимущественно ацетон и бутиловый спирт. При ацетоно-бутиловом Б. сбраживаются моносахариды, дисахариды и полисахариды - крахмал, инсулин, но не сбраживаются клетчатка и гемицеллюлоза. Ацетоно-бутиловое Б. использовалось для промышленного получения бутилового спирта и ацетона, применяемых в химической и лакокрасочной промышленности (см. также
Ацетоно-бутиловое брожение и
Ацетоно-этиловое брожение).
Сбраживание белков. Некоторые бактерии из рода Clostridium - гнилостные
Анаэробы - способны сбраживать не только углеводы, но и аминокислоты. Эти бактерии более приспособлены к использованию белков, расщепляемых ими при помощи протеолитических ферментов до аминокислот, которые затем подвергаются Б. Процесс сбраживания белков имеет значение в круговороте веществ в природе (см.
Гниение).
Пропионовокислое Б. Основные продукты пропионовокислого Б., вызываемого несколькими видами бактерий из рода Propionibacterium, - пропионовая (CH3CH2OH) и уксусная кислоты и CO2. Химизм пропионовокислого Б. сильно изменяется в зависимости от условий. Это, по-видимому, объясняется способностью пропионовых бактерий перестраивать обмен веществ, например в зависимости от аэрации. При доступе кислорода они ведут окислительный процесс, а в его отсутствии расщепляют гексозы путём Б. Пропионовые бактерии способны фиксировать CO2, при этом из пировиноградной к-ты и CO2 образуется щавелевоуксусная к-та, превращающаяся в янтарную к-ту, из которой декарбоксилированием образуется пропионовая к-та:
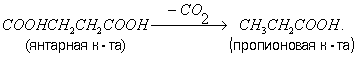
Существуют Б., которые сопровождаются и восстановительными процессами. Примером такого "окислительного" Б. служит лимоннокислое Б. Многие плесневые грибы сбраживают сахара с образованием лимонной кислоты. Наиболее активные штаммы Aspergillus niger превращают до 90\% потребленного сахара в лимонную кислоту. Значительная часть лимонной кислоты, используемой в пищевой промышленности, производится микробиологическим путём - глубинным и поверхностным культивированием плесневых грибов.
Иногда по традиции и чисто окислительные процессы, осуществляемые микроорганизмами, называется Б. Примерами таких процессов могут служить уксуснокислое и глюконовокислое Б.
Уксуснокислое Б. Бактерии, относящиеся к роду Acetobacter, окисляют этиловый спирт в уксусную кислоту в соответствии с суммарной реакцией:
Промежуточное соединение при окислении спирта в уксусную кислоту - уксусный альдегид. Многие уксуснокислые бактерии, кроме окисления спирта в уксусную кислоту, осуществляют окисление глюкозы в глюконовую и кетоглюконовую кислоты.
Глюконовокислое Б. осуществляют и некоторые плесневые грибы, способные окислять альдегидную группу глюкозы, превращая последнюю в глюконовую кислоту:
Кальциевая соль глюконовой кислоты служит хорошим источником кальция для людей и животных.
Лит.: Шапошников В. Н., Техническая микробиология, М., 1948; Прескот С., Дан С., Техническая микробиология, пер. с англ., М., 1952; Пастер Л., Избр. труды, пер. с франц., т. 1-2, М., 1960; Кретович В. Л., Основы биохимии растений, 4 изд., М., 1964; Фробишер М., Основы микробиологии, пер. с англ., М., 1965; Фердман Д. Л., биохимия, М., 1966; Работнова И. Л., Общая микробиология, М., 1966.
В. И. Любимов.
Схема спиртового брожения.



.svg?width=200 "Общая схема спиртового брожения")

.svg?width=200 "Схема пропионовокислого брожения")



